CRISPRs(Clustered Regularly Interspersed Short Palindromatic Repeats) function as a defense system, which protects bacteria and archaea agains mobile genetic elements (such as phages or plasmids). In contrast to other gene transfer and page defence mechanisms, CRISPR interference is an adaptive immune system. This acquired immunity is obtaine via integration of new CRISPR spacers. The degradation of the mobile genetic elements is mediated via an specific interaction of a small RNA (crRNA) and the target nucleic acid (DNA or in some cases RNA). This immunity can be also passed to offsprings (Lamarckisms ) (5). It can be reprogrammed to destruct an invading DNA which the bacteria have not previously encountered. CRISPERs are found in approximately 40% sequenced bacterial genomes and in 90% sequenced archaeal genomes (Jansen et al., 2002) (3). The presence or absence of CRISPRs is not characteristic for a particular taxon among Bacteria. The number of CRISPR loci per genome is limited, the current record is 18 clusters (archaea).
CRISPRs were first described in 1987 by Ishino and collegues, who found 14 repeats of 29 base pairs that were interspersed by 32-33bp non-repeating spacer sequences (1). In the following years much more CRISPR sequences were identified in various archael and bacterial species.
Jansen and co-workers found four CAS genes adjacent to these repeats. Additional studies detected more CAS genes in proximity of the repeats. These sets of genes are not present in bacterial species which lack CRISPRs. Based on the structure of these loci, in 2002 Jansen et al. named these repeats CRISPRs (2).
Jansen et al. (2002) identified CRISPR sequences by using NBLAST (sequence similarity program) (3). They considered the possibility that CRISPR loci might exist in many bacterial species with the characteristic motif of alternating short repeats and unique sequences. They found CRISPR motifs approximately in 50% of bacterial species. The repeat sequences were rather small: 21-37bp, the spacer sequences were of a similar size like the repeat sequences. Jansen et al. discovered that the common structural characteristics of the CRISPR loci are:
the presence of multiple short direct repeats. These repeats are usually evolutionally conserved and show no or little sequence variation
the presence of non-repetitive spacer sequences between the repeats. These spacer sequences are approximately of the same size as the repeats
the presence of a common leader sequence
the absence of long ORF within the ford focus
the presence of cas 1gene accompanied by the cas2, cas3 or cas4 genes
Mojica et al., 2005, reported that the CRISPR intervening sequences and the spacers are not as unique, as previously considered, but they are derived from preexisting sequences. They show that CRISPR spacers derive from either chromosomal or transmissible genetic elements such as bacteriophages and conjugative plasmids.
Mojica et al., 2005
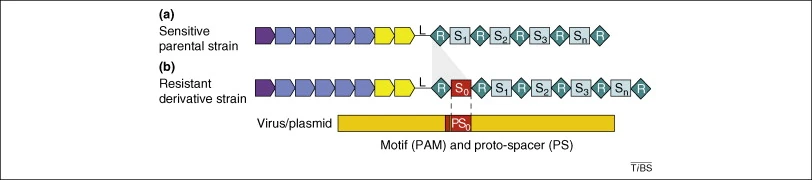
In 2007 a study by Barrangou et al. showed that the CRISPR loci function as an adaptive immune pathway that protects bacteria (Streptococcus thermophilus) against phage infection (4.). They have shown for the first time the sequence identity between the spacer in the bacteria and its match in the phage genome (the protospacer).
Barrangou et al., 2007
Barrangou et al. analyzed the CRISPR sequences of various S. thermophilus strains. They discovered that differences in the number and type of spacers determines phage sensitivity or resistance. Phage-resistant bacterial strains seemed to obtain additional spacers. To determine the origin of and function of these additional spacers Barrangou et al. have selected a phage-host model consisting of a sensitive wt S.Thermophilus strain and two phages: 858 and 2972. They selected independently nine phage-resistant mutant by exposing the Wt strains to phage 858, 2972 or simultaneously to both and analyzed the CRISPR loci of these strains. They observed that at CRISPR1 locus 1 to 4 additional spacers were inserted in a polarized manner (toward one end of the CRISPR1 locus). By sequence analysis of the additional spacers inserted in the CRISPR1 locus of the various phage resistant mutants they revealed the similarity between the sequences in bacteria and sequences found within the genomes of the phages.
Structural features[]
Marraffini and Sontheimer, 181
[]
Repeats[]
Repeats are almost always identical in size and in their sequence (1) (3). They are divergent between species, but can be clustered in at least 12 group based on their sequence similarity. Some of these groups contain 5-7bp palindromes, which form a secondary structure, a stem loop, on the repeat. Many of the repeats also have a conserved 3' terminus GAAA (C/G). This terminus and the secondary structures serve as binding sites for Cas proteins. Repeats can be from 23-50nt long, but have an average length of 31nt (3).
Spacers[]
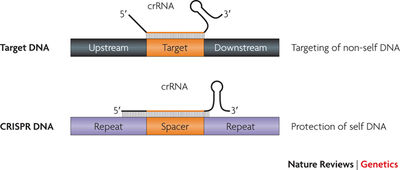
Spacers are unique sequences in the CRISPR system. It was observed that spacer sequences frequently match with high sequence identity to phages and extrachromosomal elements (1). Spacer sequences range from 17-84nt with an average of 36nt (they are therefore of a similar size like the repeats) and are functional in both sense and anti-sense orientations.
Protospacers: Protospacers are sequences present in phage genomes that are recognized and uptaken by bacteria as spacers. Protospacers seem to be evenly distributed in the phage genomes. A short motif (i.e. AGAA and GGNG) is often present in phage genomes 1-5 nucleotides downstream of protospacers. These motifs are called PAMs (Protospacer adjacent motifs) and are important for the recognition or cleavage of the phage sequences by the CRISPR system.
The number of repeat-spacer units is limited and varies from just a few units to several hundred units. The average is 66 units per cell.
Leader[]
Thel leader sequence is located at the 5' end of most CRISPR loci (1). It is long up to 550bp and is usually AT rich. Leaders (as well as repeats) lack an ORF and are not conserved. A new repeat-spacer sequence is always added between the leader and the previous repeat-spacer unit. It is possible, that the leader also acts as the promoter.
CAS genes[]
CAS genes (CRISPR associated genes) are sets of gene families that associate with CRISPRs (1). More than 40 cas genes have been identified up to date. The most common member of the Cas gene family is cas1, which is up to date linked to almost all CRISPR systems and serves as a universal marker. Cas protein have special domains with endonuclease activity, helicase activity, RNA and DNA binding activity and domains, which are involved in transcription regulation. Cas proteins also cleave pre-crRNA precursors at a specific site in the repeat sequences and therefore form a mature crRNA.
Sorek et al., 2008
Function and mechanisms of CRISPR interference[]
There are three distinct stages in the CRISPR defense mechanisms (5):
Oost et al., 2009
Adaptation of the CRISPR via integration of short sequences of the invaders as spacers[]
In this phase CRISPR loci incorporate spacers to programme their activity agains invading elements such as plasmids or phages. This allows the cell to adapt to the invadors from the environment. The acquisitions of new spacers occur at the leader site (5') of the CRISPR. CRISPRs can therefore serve as a record of previous encounters with mobile genetic elements.
Expression of CRISPRs and subsequent processing to small guide RNAs[]
Transcripts of the whole CRISPR regions are called pre-crRNA, these are processed to small crRNAs. The CRISPR processing is mediated by eight genes: Cas1 (integrase/nuclease), Cas2 (nuclease) and Cas3 (helicase/nuclease) and by a Cascade set of genes/proteins (5). Cascade genes include Cse1-Cse2-Cse4-Cse5-Cse3 (CasABCDE) and are variable. The CRISPR transcript forms a stable interaction with the Cascade complex. The protein CasE catalyzes the specific processing of the transcript to small mature crRNAs with a 8-9nt repeat fragment at the 5 'end and a longer and more heterogenous repeat fragment at the 3' end. These repeat sequences present at both ends of the spacer sequence prevent the cell from splicing its own DNA.
Marrafini et al., 2010
Interference of target DNA by the crRNA guides[]
The mature crRNAs in a complex with Cas proteins have a base-pairing potential, they serve as guides and recognize the invasive traget (2). The crRNA sequence has to be identical with the invading sequence, since even a single mismatch compromises CRISPR interference. The processed CRISPR spacers therefore function as the microbial analogues of siRNAs: they bind to a RISC-like complex with cas proteins, recognize the DNA/RNA that is expressed from the foreign element, which results in degradation of the mRNA by other Cas proteins (cas3) (1).
CRISPERs and evolution of bacteriophages[]
Phages can overcome CRISPR immunity by acquiring a single mutation in or around a target sequence. There must be a perfect match between the crRNA and the phage RNA for interference. In natural environments, the host cell is exposed to many phage variants. These phage variants therefore recombine to counteract CRISPR immunity.
{kind=link}

{kind=link}

{kind=link}



{kind=link}


{kind=link}


{kind=link}

{kind=link}